Intro l
Sensation l
Revolution l
NeuroPhilo
Epilogue l
Biblio
Neurophilosophical Inquiries
©
Wim van den Dungen
these inquiries have been updated in subsequent
books :
Regulae I (2025) and
Regulae II (2025)
Bits of Neurology for Philosophers
1
Remarks about Intelligent
Design.
2
Temporal-limbic versus fontal lobe spirituality.
3 The mammalian brain,
the democratic neuron and philogenesis.
4 Major brain structures and their functions.
5
The triune brain : reptilian, mammalian and human.
1 Remarks about Intelligent Design.
Dembski & Behe (1998) and
Hamilton
(2002) argue the universe shows intelligent design. Central to
the debate is the question whether the organization of the universe in general
and the emergence of life in particular is a matter of accident ?
Hoyle
(1986)
concludes random events and change occurrences are insufficient to account
for the complexity of living organisms. Earlier,
Prigogine (1979) saw
the return of finality in open, dissipative systems.
Instead of blind chance, the universe shows "finality". This
"causa finalis" of old (cf. Aristotles, Metaphysics, Book 1) is
reformulated as the "anthropic principle", stating that if any of
the physical constants were to vary from the fine-tuned values we determined,
life would not be possible. Its "weak" version reasons
certain cosmological features are conductive to a universe tuned to life,
whereas the "strong" version holds the universe to be bound to
produce conscious and intelligent beings. This strong "tuning"
suggests a Great Architect, a principle ordering the parameters of the
universe and its life. Every slight variation in the known natural constants,
drastically alters the organization of the universe. Of all possible universes
(cf. Leibniz), ours allows for life and consciousness. The universe was meant to
be conscious and humanity allows it to be so. These arguments are studied in
more detail in
Does
the Divine exist ?, 2005.
In biology, neo-Darwinism is also challenged. Indeed, the notion
of "variation" as "random mutation" is consistent with the
denial of purposeful design, making natural patterns without inherent plan (Monod,
1970). Recently, a progressive metamorphosis, with the emergence of
increasingly complex and intelligent species in a step-wise, sequential pattern
was proposed (Joseph, 2002). Indeed, large-scale protein innovation
(Aravind,
2001), so-called "silent genes" (Henikoff, 1986,
Watson, 1992), the
precise regulatory control of genome novelty (Courseaux &
Nahon, 2001) and
the overall genetically predetermined "molecular clockwise" fashion of
the unfoldment of the human being (Denton, 1998),
etc. underline
the evolutionary
metamorphosis theory of life and intelligent design (weak anthropic principle).
According to Darwin's evolutionary theory, phenotypic variation
originates from random mutations independent of any outside selective
pressure.
"However, recent findings suggest organisms have evolved
mechanisms to influence the timing or genomic location of heritable
variability. Hypervariable contingency loci and epigenetic switches increase
the variability of specific phenotypes ; error-prone DNA replicases produce
bursts of variability in times of stress. Interestingly, these mechanisms
seem to tune the variability of a given phenotype to match the variability
of the acting selective pressure. Although these observations do not
undermine Darwin’s theory, they suggest that selection and variability are
less independent than once thought."
Rando, O.J. & Verstrepen, K.J. : "Timescales of
Genetic and Epigenetic Inheritance", in Cell, vol 128, 655-668, 23,
2007, my italics.
Intelligent design calls for a Designer,
conceptualized as :
Pantheism is difficult to
maintain, for the characteristics of the
Designer are so different from those of the design itself. It downgrades the Designer
(reduced to a "subtle" kind of matter or "pneuma") and so
harms the principle of intelligent design itself. Nevertheless, pantheism is the
most intelligent option in an immanent metaphysics, positing the Divine
insofar as it is immanent and constantly present everywhere in the world
(cf. the argument of conservation).
Theism is a consequent view
on the Designer (supposed to be radically different from the design itself), but
then it become difficult to understand how the design can come into
being if its Designer is so far removed from the design. A solution through mediation (a "Pharaoh", a "Buddha", a "Christ")
mystifies the nature of the mediator and cancels the procedure, for nobody
understands how the Designer designes the mediator, except dogmatically. The notion
everything is part of the Designer (cf. Qabalah, were creation happens in
a space vacated by God), is a stronger position, although the
conceptualization of the Designer remains paradoxical (but this does not affect
the design). How can what is "far" (transcendent) simultaneously be
"near" (cf.
Porete's "Loing-Près") ? The paradoxical
bi-polarity of the Divine, the
theonomy of perplexity
of the mystics is however in accord with this position.
Furthermore, transcendent metaphysics always exceeds its limitations, as
nondual thought is not conceptual but of the nature of direct discovery
and realization (cf.
Intelligent Wisdom, 2007).
Pan-en-theism, as in Qabalah, seeks to find the "traces" of the Designer in the
design. These enable us to remodel (perfect) parts of the design in the light
of what is known about how the Designer designed the design. The latter is an
integral part of the Designer, not some entity "outside". The Designer
has no "outside", for the unlimited has no borders. The design is
positive (full) existence within the negative (empty) being of the
Designer, although this "emptiness" is rather a potential, virtual
reality or set of all possibilities than an absolute absence of anything
(an enduring nothingness).
Separated from the universe, there is
virtuality, potentiality and possibility, which is more than non-entity. That
something is the "prima materia" or transcendent source of
being.
This negative, virtual existence of the Designer is Designer consciousness
(i.e. awareness untained by matter or information). Designer consciousness (the absolute in its
absoluteness) creates an intelligent design, i.e. a finality expressing the
eschatological activity of the Designer. This happens by confining a spark of this sheer
intentional, preexistent Being within certain limits. This allows for
organization, composed of consciousness, information and matter. This virtual,
preexistent, ineffable, primordial, pre-creational, limitless, infinite Being contracts the manifestation of the design within the sphere of its
own set of "natural" laws. As before creation nothing is manifest, the
Designer is immaterial. As before creation no differentiations exists, the
Designer is not informational (the names of the absolute are part of the design,
not the Designer). This creativity is exclusively intentional and
untouched by, unmingled with, veiled from the processes of involution &
evolution.
Only such preexistent, pristine consciousness is able to
create its own intentional objects, to wit : the design of universes,
operated by consciousness, information and matter. As the consciousness at work
within a universe is always limitated by its co-relative bond with information
and matter, i.e. the operant code of existence ruling the logical confines of
this universe and its physical conditions (spatiotemporal, dynamical, atomic),
only the Designer is deemed to be absolutely free.
Generation
initiates a universe ("exidus"). With the manifestation of life, consciousness
eventuates and the return-sequence is initiated
("reditus"). This evolutionary sequence (matter, information,
consciousness) initiates the progressive metamorphosis, step-wise producing
increasingly complexer creatures, able to manifest, execute & compute more
meaningful intentional states. Both sequences are dynamical, fluent, creative
& intelligent. With the emergence of the human brain, the ability to
materialize consciousness dramatically altered the natural sequence of events on
this planet, showing that the intentional world is indeed the realm of
freedom.
In this ontological speculation, the Designer is present in the fundamental
intelligence of the material design ("arché") and the meaningful
complexification of its elements ("telos") of our universe. This is
the text, script, scenario (information) the Designer devised. This "presence" of
the Designer as "law" rules the manifestation of consciousness and the
spiritualization of matter. From a metaphysical perspective, science is then the
discovery of the abstract trace of the Designer in the design.
So in Western creational ontology, the process of involution/evolution, the
life-span of this world, is able to run its course and
complete itself as a result of three fundamental eschatological impulses :
-
material impulse :
the Designer as "Creator" fashions a material matrix as an
object within the limitless full-emptiness of pre-creation (cf. the
"Nun" in
Ancient Egypt or "Ain
Soph" and "tzimtzum" in
Qabalah). An
infinite being, a pure act (or unmoved mover - cf. Aristotle) restricts
itself to create a real but finite being. This material matrix, or "hardware" of
our universe, is a unity of chemical elements (especially helium, hydrogen),
a host of elemental particles (like the photon), as well as a series of physical forces (superforce, weak, strong,
gravity, electro-magnetism) and their differences (or energy, conceived as
the product of differences). For Hegel, the formidable extention of the physical universe speaks of the
intensity of
the non-spatial spirit (or the names, signs, traces, residuals, "reshimu" of the Designer
present in the design) ;
-
informational
impulse : the Designer as "Great Architect", organizes the material matrix
through physical constants and intelligent laws reducing all possible cosmoi
(all variations of the
components of the matrix) to our present, observable universe, with its characteristic universal laws of
physics, open windows to the emergence of life. Fundamental is the universal repetitive cycle of expansion and
contraction of all matter within this universe and maybe between universes
(waning & waxing forever). Code is added to the
material matrix, allowing for the formation of DNA, the informational
building block of life and storehouse of cellular intelligence, memory,
planning skills & language. The vital principle holds : our universe
is built to eventually produce the evolution of living beings gifted with intelligence, or the "software" of this universe ;
-
consciousness
impulse : the Designer as "Great Artist", replicates in miniature the proto-type of the universe
(macrocosmos) as an "Ultimate Concern" (cf.
Tillich,
1952) or primordial example aspired to by creatures (microcosmos) able
and invited (not forced) to follow the "straight path" (the
convenant between humanity and the Designer). Hopefully this leads to a perfected seering
and meaning capacity in this universe (cf. "Omega" in
Teilhard de
Chardin, 1959, or nondual thought - cf.
Intelligent Wisdom, 2007). In doing so, the universe
changes for the sake of the
endless meaning-complexification arrived at by highly complex beings such
as humans. The anthropic principle holds : the material and informational
processes of the universe have parameters which favour the emergence of
hominoids (pre-hominids) and hominids. They have brains able to process (compute,
transmit, mediate) consciousness and volition, or the "userware"
of this universe.
Four analogies provide a strong backing for the case
or premiss presenting the
non-spontaneous becoming of the actual world process.
How to detect non-spontaneous "design" ?
-
design by
analogy of human products :
the proximate cause proportional to the order, harmony, fitness &
freedom observed in the world can be identified (named) by following the
analogy of products of human design. In doing so, only the "form" aspect
of the world is observed to identify design. In this way, the "matter",
or substance of the world, is not targeted, and it is no longer
necessary to prove in addition, that the things of the world, given the
laws of nature, were in themselves incapable of such order and harmony.
Hence, to avoid backing the premiss, it is accepted that no supreme
intelligence exists in the material substance of the things of the
world. In the traditional Peripatetic account, four causes are at work
in the world : material, efficient, formal & final. By analogy of human
products, the design involves the formal and final causes only ;
-
design by
analogy of outcomes in living organisms : all living things seem
tailor-made for their function and appear to interact purpose-fully with
their environments : animals use camouflage, most parts of our bodies,
down to our DNA helix, are very delicately engineered, and large numbers
of apparent coincidences exist between various living organisms, etc.
These highly ordered biological schemata seem places of reference
to back the premiss, for how could such a complexity rise out of
simplicity without a pattern of intelligent choices ? The chances are
small enough, given what science demands in other areas, to dismiss
spontaneous, random activity. Nevertheless, this study of outcomes was
seriously affected by the discovery of the Darwinian principle making
organisms evolve by natural selection, adaptations and (random)
mutations. If all biological events can be explained by this
principle (turned into a paradigm), then indeed there is no "purpose"
behind the grand natural symphony. Darwin (1809 - 1882) and
neo-Darwinism were able to explain much of the data of his time and the
first half of the previous century. Even societies could be studied in
terms of the survival of the fittest
(Monod,
1970). But, recent studies show how the
theory has been unable to account for certain more subtle phenomena
uncovered by the biochemistry of the last 50 years, mostly related to
complex events such as protein transport, blood clotting, closed
circular DNA, electron transport, photosynthesis etc. So, beyond the grip of Darwin's macroscopic view,
on those more subtle levels of biology and biochemistry, design may be
detected and purposeful arrangement of parts suspected. A revised
analogy of subtle outcomes becomes thus again possible, leading to a
more comprehensive backing of the premiss ;
-
design by
analogy of the forms of the laws of nature : Maxwell (1831 -
1879) pointed to molecules as entities not subject to selection,
adaptation & mutation. The contrast between the evolution of species,
featuring biological changeability, and the existence of identical
building blocks for all observed actual physical entities is crucial. Calculate
the odds of spontaneous emergence, given the effectiveness of Newton's laws on the mesolevel (the
inverse-square law of gravity being optimal for the becoming of the
Solar system), our knowledge of what happens in stars (in particular the
production of carbon and oxygen) and the cosmology of the Big Bang. A choice has to be made
between either an intelligent design (which does not offend except the
ill informed) or a monstrous random and blind sequence of accidents
producing a gigantic complexity, in other words either a natural higher
intelligence or the ongoing mathematical miracles of a blind nature
morte. Indeed, ad contrario, the form of the laws of nature underlines the presence of a deep-laid scheme, representing an
accurate mathematical descriptions of the natural order (both in genesis
as in effect). Although no "consensus omnium" has been reached,
the laws of nature likely accommodate biology ;
-
design by
analogy of fundamental constants : the actual irreducible
mathematical presence of immutable natural building blocks such as the
natural constants, gives a palpable proof of the existence of something
independent of every human measurement (and its biological
constitutive). These constants define the fabric of physical reality
and determine the nature of light, electricity and gravity. They make
particles come into existence and fundamental forces work. They
actualize the laws of physics by giving equations numerical quantity and
are necessary in the logic of physics. What can be said about the
particular values takes by these constants ? The conditions for order
and eventually life to develop have been found to heavily depend upon
these constants. Indeed, although mathematically, the equations of
physics, representing the fundamental architecture of the order of the
world, also produce outcomes when other quantities of the same constants
are put in, the world would be lifeless and barren (instead of a
haven for incredible complexity) if even a small amount of these values
would be changed. Ergo, the various values of the constants of
nature were designed, and pre-planned. An infinite number of different
worlds are possible, but only in one are order, fitness, beauty and life
actual. Only our universe has observers witnessing it.
Instead of blind chance, the universe
has "finality", i.e. an ultimate aim or purpose. This
"causa finalis" is
the notion backing the "anthropic principle". For if any of
the natural constants were to vary from the fine-tuned values physics determined,
life as we know it would not be possible. This "weak"
anthropic principle posits cosmological features conductive to a universe tuned to
and generative of life as we know it. Accepting life is bound to be
observed, the "strong" principle affirms the universe is bound to
produce conscious and intelligent beings. This addition of the
observer or witness is a demand of quantum theory. The observer is thus always part of the experiment. The strong
version argues for an immanent Architect of the world (explaining the
unity of the world).
-
weak
anthropic principle : the fabric of the world is conductive to
life ;
-
strong
anthropic principle : the fabric of the world is conductive to
the observation of life and the continuum of all observations imply an
immanent Witness of all possible forms of life.
The order of the world
proposed by science is no longer Newtonian, although most equations of
relativity can be made "classical" by eliminating the
Lorenz-contractions accompanying high speeds. To solve the equations
covering most practical matters
at the mesolevel of the macroscopic, the Euclidian and Newtonian notions
about reality are adequate. But deep down, at the microlevel of physical
reality, in the vast so-called empty spaces between electron and atom
core and within the core itself, potentialities and propensities exist
which are ruled by a different set of laws.
Besides the strange logic at work in classical quantum mechanics, the
more "revolutionary" zero-point physics, or
free energy physics (Puthoff, 1989), understands the vacuum of
space as a "plenum", i.e. a fullness of energetic potentialities in
balance. This equilibrium prevents the enormous energy potentials from
becoming actual, which therefore appears as a void or a vacuum. But
every point in empty space, is a locus of convergence of humongous
energies, coming from all directions simultaneously, balancing them out. At any
point where there is an imbalance or asymmetry in this omnidirectional
canceling of energies, there appears a disturbance known as matter. All
particles of quantum physics are various modes of asymmetry of the
zero-point field of the vacuum with itself. This zero-point energy is
seen as the result of the unpredictable random fluctuations, which, in
classical theory, are all zero. But, even at a temperature of absolute
zero, where no thermal agitation can have effects, the flux remains.
2
Temporal-limbic versus fontal lobe
spirituality.
Let us consider these landmarks :
observable universe ;
ca.5 billion years ago : formation of our Sun ;
ca.4.6 billion years ago : the formation
of the Earth ;
ca.3.5 billion years ago : Sun-worshipping single-celled organisms
;
ca.3 b illion
years ago : the ability to exchange information between members of the
animal kingdom ;
ca.1 billion years ago : sex becomes a
widespread form of reproduction in oxygen-breathing, multicellular creatures
;
ca.700 million years
ago : creation of the first neuron (one without axons and dendrites) ;
ca.500 million years ago :
first primitive aggregates of neural ganglia into lobes ;
ca.500 million years ago :
start of formation of primitive limbic system ;
ca.500 million years ago :
first vertebrates swim the ocean and plants invade land ;
ca.300 million years ago :
first great forests ;
ca.100 million years ago :
first mammals appear ;
ca.30 million years ago :
first apes ;
ca.5 million years ago
: first erect hominids emerge and spread across the face of the planet ;
ca.2.5 million years ago :
earliest samples of chipped pebbles, the work of Homo habilis ;
ca.1.6 million years ago : Homo
erectus walks and uses the same stone tools for a million years ;
ca.450.000 years ago :
limbic system functional in sharks ;
ca.500.000 years ago : brain
of hominids doubled in size ;
ca.500.000 years ago :
emergence of the Homo sapiens neanderthalensis ;
ca.100.000 - 40.000 years
ago : the Cro-Magnon brain is formed (Homo Sapiens sapiens) ;
ca.40.000 - 10.000 years ago : the human brain eventuates
;
ca.10.000 BCE : start of the Neolithic and
human civilization .
By 30 million years, higher
primates had emerged and dwelled in Africa, China and
India. From these wide ranging hominoid stocks, a variety of hominoid
pre-hominids began to evolve : Dryopithecus, Sivapithecus, Ramapithecus,
Ankarapithecus, Ouranopithecus, Giganotopithecus, etc ... These
protohuman
types
experienced the end of the receding forests and the expansion of the savanna &
grassland. As a result, around 5 mllion years ago, some hominoids began to
increasingly live upon the ground.
"... our feet and legs are the most human things
about us and are ultimately answerable for what happened late in our
evolution. For one thing, standing erect on our legs freed out hands from any
work in moving us about."
Howells, 1997, p.55.
Living on the ground caused further
evolutionary metamorphosis and gave rise to the first hominids, such as
Australopithecus Afarensis (Joseph, 2002),
who's erectness is clear (Howells, 1997, p.81). A wide range of Australopithecus
emerged 5 million years ago, throughout Africa as well as China and Java. Around 2 to 3 million years ago, Australopithecus was
joinded by the Homo habilis ("able with hands") of which several varieties appeared in Africa, China
and Indonesia (Howells, 1997, pp.93-120).
Austrolopithecus and Homo habilis continued to evolve giving
rise
to a wide variety of species, collectively referred to as the Homo erectus, who appeared ca.1.6 million years
ago. A critical brain expansion had happened :
-
Australopithecus
: 450 - 520 cc, who roughly had our teeth and feet ;
-
Homo habilis :
600 - 800 cc, who made simple stone tools ;
-
Homo erectus : 900 - 1000 cc, larger brain and a skeleton like our own ;
-
Homo
Neanderthalensis : 1.033 - 1.681 cc, larger brain but frontal
lobes less developed ;
-
Homo Sapiens
sapiens (Cro-Magnon) : 1.600 - 1.681 cc, large brain with
developed frontal lobe ;
-
Homo Sapiens sapiens
(present-day humans) : ca. 1450 cc.
"It must be more
than a coincidence that tools and the early Homo appeared at the same
time. Brains continued to expand, and tools continued to become more varied
and better defined."
Howells, 1997,
p.100.
With the Homo
erectus the brain gradually doubled in size. He emerged in
Asia, Africa and Europe. Homo erectus harnassed fire (to keep warm, to provide
light and to cook their food), developed
crude shelters and home bases and utilized earth pigments (Joseph,
2002, p.118). But in a million years he never
changed his tools ...
Archaic Homo sapiens neanderthalensis (or Neanderthals) first emerged almost 500.000 years
ago. Until 300.000 BCE, Homo erectus continued to dominate parts of the planet,
and so Neanderthals and Homo erectus shared the planet in different geological
locations for almost 200.000 years. Ca.100.000 BCE, the Homo sapiens sapiens
(Cro-Magnon) appeared in North and South Africa and between 50.000 and 40.000
BCE, they spread upward into the Middle and Far East, then into Europe, China,
the Americas and Australia (Joseph, 1993, p.29).
With them, we've reached our true ancestors.
"... let us be warned that the Upper Paleolithic
Europeans have no copyright on the apparent 'sapiens explosion'. It would
appear instead that the European Upper Paleolithic was more a ripening
expression of the new human level, no a cause of it, and we need much more
evidence on the nature of this florescence."
Howells, 1997,
p.185.
Different species of hominids
were living in different parts of the world
simultaneously :
-
Homo erectus :
flourished from 1.6
million years until 27.000 BCE (Java) ;-
Homo
sapiens neanderthalensis
: from 500.000 until 29.000 BCE ;-
Homo Sapiens sapiens : from
100.000 until 10.000 BCE.
"Hence,
similar to the step-wise worldwide pattern of multi-regional, multi-phylectic
metamorphosis which has characterized the progressive emergence and increased
complexity of plants and animals, the available evidence suggests that human
'evolution' has unfolded multi-regionally in a step-wise, progressive fashion,
with some groups lagging far behind and others being left behind altogether and
becoming extinct."
Joseph, 2002, p.121.
The differences between Homo sapiens neanderthalensis (culminating
in the Middle Paleolithic, i.e. between 150.000 and 35.000 BCE) and Cro-Magnon
Homo sapiens sapiens (of the
Upper Paleolithic, i.e. between 35.000 and 10.000 BCE)
reveal a major point of interest for neurospirituality (Joseph,
1999, 2002).
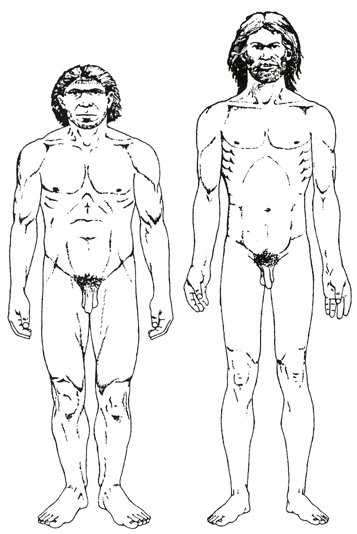
Neantherthals had a sloped and stunted frontal cranium, whereas the Cro-Magnon
had a brain larger than that of the average contemporary human being.
In 1868, during construction for a railroad, in a shallow cave at Cro-Magnon in the Dordogne area of
southern France, a number of obviously ancient human skeletons were found. The
cave was investigated by the French geologist Louis
Lartet, who uncovered five archaeological layers. The human bones found in
the topmost layer proved to be between 10.000 and 35.000 years old. Cro-Magnon was robustly built and powerful and is presumed to
have been about 166 to 171 cm (about 5 feet 5 inches to 5 feet 7 inches) tall.
The features were generally heavy and solid. The forehead was straight, with
slight browridges, the skull long and narrow, and the face short and wide. The
ape-like features still apparent in the Neanderthals had vanished.
Let us highlight a few important differences :
hominid left no physical traces of ritualized
behavior. The notion they were the first living creature on Earth able
to perceive a spiritual reality, and this because their complex brain
contained all the key neuronal structures for creating language -cf. the
parietal lobe- (Newberg, d'Aquili & Raus,
2002, p.66) is a controversial idea.
Cranial analysis suggests the opposite (cf. infra). In a million years, Homo
erectus was unable to invent new tools. They
lived in small groups, inhabited caves, and finally mastered fire,
enabling them to, as it were, "domesticate" the night and sleep
without fear (and invent the waking dream ?) ;
Neanderthals
: they possessed a developed inferior temporal lobe. Together with the
limbic structures within it, these neuronal networks allowed them to execute
individual, emotional and religious experiences, including long term emotional
attachments and intense love (Gloor, 1997). But with such a brain, they were
unable to make complex tools or compute abstract thoughts. They were
people of passion. Nevertheless, it seems the experience of Divine
Presence was already part of their existence and behavioral patterns, making
of the Neanderthals the first hominids with spiritual capacities. Some
authors have argued the evidence of this is sparse and controversial.
But general agreement exist about the fact that due to their
deficient prefrontal cortical development, the Neanderthals -compared to
modern humans (Balter,
2002)- lacked symbolic charge. Their tools were "use-specific" and served a
unidimensional purpose ;
Cro-Magnon
: with the increase of the frontal lobe, a "symbolic
explosion" (Leroi-Gourhan, 1982) could take place. The Cro-Magnon tool
kit consisted of well over 125 items. These "intelligent" humans
were the first "intellectual giants", producing artists,
musicians, craftsmen, sorcerers (shamans), priests, hunters, fishermen,
gatherers & herbalists. Some were city dwellers, others lived in tents
made out of animal skins. They had complex mortuary practices. The role of
the mother goddess was omnipresent (a practice
to continue well into the Neolithic - cf. the great goddess in Predynastic
Egypt). She was attended by animals and shamans.
They invented initiatoric rites, in which one had to enter the hidden
entrance of an underground cave and crawl a considerable distance, through a
twisting, narrrowing, pitch black tunnel before reaching an underground
cathedral lit with light candles and lamps, with painted animals on its
walls. They were the first to use such devices to invoke the experience of
death and the subsequent initiation into a new state of consciousness. These
superior hominids were able to symbolize their religious experiences, and
thus shape religions, spiritual traditions and develop theological notions
like heaven, hell, god and goddess, as well as shamanism and priesthood.
Neanderthal and Cro-Magnon
reconstructed by
Howells,
1997, p.145.
The distinction between
Neanderthal and Cro-Magnon spirituality is on record. From an evolutionary,
philogenetical perspective, the human brain emerged two fundamental executant
levels to process spiritual experience :
-
a temporal-limbic
"root" spirituality, involving a direct, emotional, awe-strikken, passionate,
profound spiritual experience and
-
a prefrontal cortex
"symbolic" spirituality,
allowing for the computation of artistic, initiatoric,
theological and abstract cultural superstructures (mediations,
stabilizations) of such experiences.
The Cro-Magnon brain was
significantly larger than the Neanderthal brain. The differential evolution of
the frontal lobe and angular gyrus is based on the study of tools, hunting
technology, artistic and symbolic developent and social organization in the
Middle and Upper Paleolithic (Joseph,
1993, chapter 6).
Spiritual experiences processed by the
inferior temporal lobe, amygdala and hippocampus focus on an individual's
profound link with a higher presence : the direct experience of what is
spiritually significant. The radical confrontation with radical otherness
can not be explained or understood. There is no elaborated symbolic
representation at this level of direct, mystical contact. The Neanderthals show it is possible to be very religious but not smart, i.e. unable to
"explain" (symbolize) and thus made decontextualized use of what has been
experienced. Nevertheless, the hereafter is sensed, as well as the importance of
death as a passage (the idea they only buried their dead to deal with odor
seems far fetched -
Rudavski, 1991).
"Neurophysiologist
Rodney Holmes, looking at the record of archaeology, theorizes that hominids
since the Neanderthals have been Homo religiosus - creatures in search
of meaning and significance."
Albright &
Ashbrook, 2001, p.156.
The spirituality executed by these temporal & limbic wirings deals with the
direct experience devoid of abstract thought and symbolizing. Volitional
manipulation is not at hand. The software of the "God-spot" runs on the same
neuronal structures computing animal emotions and storinge highly
individual
experiences. Terror, rage, violence, emotional arousal, pleasure, joy, trance,
ecstacy and the
holy cohabitate ! These amygdalean associations are not taken away with the
development of frontal lobes, but integrated in a higher structure, enabling a
more symbolized, linguistic and artistic approach of spirituality, inventing
rules and codes to "channel" the "basic" Neanderthal sensing
of the "real" transmitted by the limbic system.
"Adam, the being created with two hands, is a locus
of coincidences of opposites."
Sells,
1994, p.86.
With the emergence of the frontal lobes, a new feature was added to the
existing temporal executives. A "pure" frontal lobe spirituality never
existed, nor will it. The frontal lobes enabled the hominids to symbolize the experience of
the sacred, higher presence (in language), and so an element of manipulation and spiritual
craftsmanship entered into the picture together with self-awareness (a conscious
division between the experiencing "I" and the object of
spirituality, i.e. radical otherness).
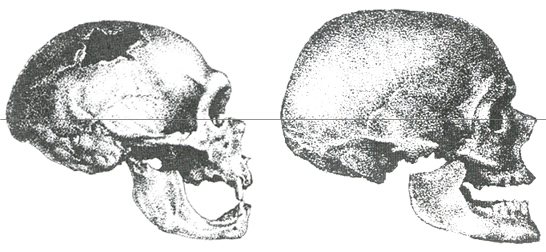
Neanderthal and Cro-Magnon
craniums
Joseph,
2002, p.349.
Between ca.35.000 - 29.000
BCE, Neanderthals and evolved Cro-Magnon shared the Earth (ca. 10.000 BCE, Cro-Magnon
gave way to the Neolithic peoples). Maybe the latter exterminated the former (Joseph,
2002, p.349). The Cro-Magnon's superiority was based on abstract thought,
individuality, speech and an enhanced regulation of emotion and action as well
as tool-making and the consolidation of cultural forms. In Neanderthal
craniums, the importance of visual input and also a less developed frontal
lobe stand out. The receding eyes of the Cro-Magnon made room for a frontal
development, which was added to the overall development of the cerebrum. If
the Neanderthal did not possess an angular gyrus (cf. infra), as made probable
by
Joseph (2002, pp.313-360), is it then not likely
this ancestor
had only perfected the skills of the archaic hominids (like Homo erectus) ? His mode of
thought was mythical and iconical. Limbic executants are at work and rule the game.
On the one hand, the frontal lobes temper the passions of temporal-limbic
spirituality by symbolizing fear and joy, as well as developing vocabularies to
give "meaning" to Divine Presence and the holy. On the other hand, too
much superstructures or a rejection of religion (as in atheism), may cause the
communication between the "old" systems and the "new" cortex
to be more unreliable or disrupted, leading to a lower level of integration of the
brain. This evolutionary block puts limitations on its executant capabilities (cf. the
absence of creativity and inventivity, as well as uplifting, joyous, profound,
holy experiences).
The symbolization of the holy experiences computed by the
amygdalean-hippocampal "God-spot", gives to their raw directness an
opportunity "to leave the limbic system" and recieve a more definitive form. Ex
hypothesi and from a neurospiritual point of view, a superstructure is
efficient if it does not disrupt or block the feedback from the prefrontal cortex to the
limbic "God-spot". In other words, symbolization is inefficient, when
it disables the use of the "God-spot" beyond the limbic system, or
condemns it to the unconscious, as in atheist ideologies. The experience of
Divine Presence has specific neuronal executants and so the mind is called to
position these experiences within the conscious field of operation of the first
person phenomenology.
In mature mystics, the profound & symbolized experience of the holy is continuous, implying an
efficient frontal lobe/temporal-limbic integration. The "God-spot"
executes the direct experience of the Divine and the "God-circuit"
lets it run through the neocortex, especially its prefrontal area and back to the
limbic system. During the frontal lobe phase, volition becomes associated (cf.
the impact of spiritual experiences on individual morality). The overall efficiency
of this circuit is, ex hypothesi, part of the evolutionary purpose of the
brain as a potentially highly complex but integrated organ, executing a
variety of mental states, making the
essential peace possible between, as it were, the limbic Neanderthaler and the frontal
Cro-Magnon. The brain may also compute the bi-polar nature of all spiritual
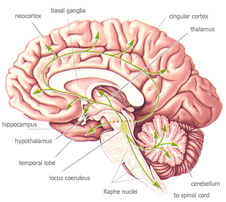
experience, rooted in the basal telencephalon (hippocampus, amygdala)
and the limbic system (thalamus, hippothalamus), but branching out towards the
neocortex, and thus enabling the expression of higher order spirituality, based on
self-experience and symbolization.
Neanderthals & Cro-Magnons show the root of
human spirituality to be buried in high-order, lower mammalian structures covered with low-order,
higher cortical networks.
3
The mammalian brain, the democratic neuron and philogenesis.
A comprehensive description of the human brain lies outside the scope of this text
and the reader is referred to
Bear, Connors & Paradiso (2001) and
other similar studies. Let me
sketch an outline and focus on those details which are of importance in our
philosophical investigation of the neuronal executants of spiritual
experience. Indeed, the human brain is the most complex piece of matter on
Earth. The description below is thus but a scaffold of some of its important
contents & functions.
general characteristics
The human brain is the most complex organ known in the observable universe.
It expends 25% of our energy intake, while consisting in only 2% of our
tissue mass.
In general, at any given moment, 1 to 10% of all
our neurons fire, on average 100 times a second. If processing speed is measured
in "FLOPS" ("floating point operations per seconds" or
the number of calculations with a floating point done in one second), and each
firing is one FLOP, then 1 teraflops is the upper limit of the
brain's information processing speed (1 megaflop = 1 million FLOPS, 1 gigaflop = 1000
megaflops, 1 teraflops = 1000 gigaflops). Counting in the synapses brings
the estimated speed up to 10 teraflops (Moravec, 1988).
The first creatures to possess one single neuron, the first cellular brains, were
able to process, store and express a billion times more information than any
sensory or motor cell could (Joseph, 1993, p.13). Living beings such as
bacteria, algae, mosses, fungi and plants have no brains, although they are
able to communicate & interact. Sponges have nervelike tissue,
concentrated around the external orifices and pores. In the nervenet of a
worm, we already recognize two large pairs of clusters of similarly
functioning neurons (neural ganglia) in the head region. Neurons are able to
plan and create memories as communicate this to other neurons in the form of
electrical and chemical messages.
The human nervous system follows the basic
mammalian plan, although -due to its extreme complexity- it is distorted as a
result of the selective growth of some parts, like the human neocortex. The
nervous system of all mammals has two divisions : the central nervous system
(CNS, encased in bone : brain and spinal cord) and the peripheral nervous
system (PNS). The latter encompasses all parts of the nervous system other
than the brain and the spinal cord.
Recently, neurocardiology and neurogastroenterology discovered two other
neuronal ganglia, respectively situated in the heart (and Solar
Plexus) and in the gut area.
In 1991, after years of extensive research, dr.J.Andrew Amour of
Dalhouse University in Halifax, Canada, evidenced the existence of a
functional "heart brain". This is an intricate network of neurons,
neurotransmitters, proteins and support cells allowing activity
independent from the "head brain". Many of the signals flowing out of
the heart to the brain regulate signals flowing out of the
brain to the heart, the blood vessels, glands and organs. The work of
the Laceys (1970) and others made clear neural messages from the heart
affect cortical activity, including the amygdala and the higher brain
centers.
There are at least forty thousand neurons in the heart. This is as many
as in the various subcortical areas of the brain. The heart is not
merely pumping blood, but demonstrating a complex set of interactions
between these neurons and those in the brain. The idea peripheral
autonomic ganglia function as "little brains" dates from the XVIIIth
century, and was formulated by Benigne Winslow, a Swedish anatomist
working in Paris. These neuronal networks were considered to perform
routine tasks required to maintain organ function, ensuring afferent
data arising from each organ did not flood the spinal cord and the
brain.
"Only one or two thousand nerve fibers connect the
brain to the hundred million nerve cells in the small intestine. Those
hundred million nerve cells are quite capable of carrying on nicely,
even when every one of their connections with the brain is severed ..."
Gershon, M. : The Second Brain, HarperCollins - San Francisco,
1999, p.xiv.
When describing the "wiring" or axons of the nervous system, two important
terms should be kept in mind : afferent ("carry to") and efferent
("carry from"). Axons transporting information from the PNS to the
CNS are afferents or sensoric axons (incoming data). Those emerging from
the CNS to innervate the muscles or glands are efferents or motoric axons
(outgoing (re)actions).
|
Mammalian Brain |
Central Nervous
System (CNS) |
Peripheral Nervous
System (PNS) |
| brain |
voluntary, somatic |
| spinal cord |
autonomic, visceral |
The PNS is
divided in two parts : the voluntary, somatic PNS and the autonomic
nervous system (ANS). The former contains spinal nerves innervating the skin,
the joints and the muscles. The latter consists of neurons innervating the
internal organs, blood vessels and glands (it is therefore also called the
visceral PNS).
Looking down on
the nervous system, we see it can be divided into two equal halves, the right
side of the brain and spinal cord being the mirror image of the left side. This bilateral
symmetry is valid for most structures within the nervous system, which come
in pairs along the midline (as can already be seen
in worms).
A side (lateral) view of the mammalian brain reveals four parts are
common to all mammals :
the cerebrum ("brain") : is the anterior (rostral-most) and largest part
of the brain - a top (dorsal) view reveals a split down the middle into two
cerebral hemispheres (separated by the sagittal fissure) - in general, the
left (right) hemisphere receives sensations from the right (left) side of
the body - a thin sheet of neurons, the cerebral cortex ("bark"),
lies just under the surface of the cerebrum ; the cerebellum
("little brain") : although dwarfed, in
humans it contains as
many
neurons as the whole cerebrum and is a movement control center with
extensive connections with the cerebrum and the spinal cord - in general,
the right (left) cerebellum is concerned with movements of the right (left)
side of the body ; the brain stem : the
stalk from which the cerebral hemispheres and the cerebellum sprout relays
data from the cerebrum to the spinal cord and cerebellum and from the spinal
cord and cerebellum to the cerebrum - it contains an important set of
ascending modulatory neurons controlling the wakefulness & arousal of
the whole cerebrum ; the spinal cord :
encased in the bony vertebral column and attached to the brain stem, it is
the major
conduit of all afferent and efferent signals and communicates with the body
via spinal nerves part of the PNS.
The outstanding
difference between the mammalian and the human brain being the cerebrum and its cerebral
neocortex. The total number of neurons in the average human brain is
ca.100 billion. Cerebellar granular cells contribute disproportionately to this
sum, for there are ca. 20 billion neurons in the typical human neocortex
and no less than ca. 109 billion granule cell neurons in the typical cerebellum.
Several studies have used neurostereological techniques to estimate the
total length of myelin fibers in human hemispheric white matter. Among
estimates ranging from 118.000 to 180.000 km, the figure of 135.000 km
(84.500 miles) is a reasonable figure to use as the total length of
myelinated fibers in the average human forebrain. Stereological measurement
of brains from across the human lifespan has demonstrated the neocortex
loses 31 million neurons per year in normal aging.
The mammalian brain (or "old" brain) has a
two-layered limbic cortical tissue. The ascension from primitive mammals to
primates and then humans implied the continuous accumulation of additional
cortex (sets of neurons performing sensoric, motoric and associated tasks),
curling up as it expanded, forming gyri. In the course of this
transition from primitive to more advanced mammals, this "new"
cortex expands and increasingly enshrouds the forebrain and limbic system.
Most animals lack the most recently acquired six- to seven-layered cortex
(or "new" brain"). But all primates with a considerable new
brain lack an angular gyrus, essential in the production of complex
spoken language (Joseph, 1988, 1990). This angular gyrus, as well as the
complexity of the human cortex, are
the outstanding neurological structures enabling us to differentiate Homo
sapiens sapiens from earlier hominids. In what follows, only
the human
cerebral cortex is always called "neocortex", whereas mammals acquire
"new" cortex.
|
Parameter |
Value |
| number of neurons |
ca.1009 |
| cortical
neurons |
ca.209
(*) |
| surface of neocortex |
ca.11 m² |
| connections per neuron |
ca.1000 |
| cortical synapses |
ca.240
trillion (*) |
| storage
capacity/synaps |
1 bit (1/8
byte) |
|
(*) Koch, C : Biophysics of
Computation, Oxford University Press - New York, 1999, p.87. |
As we shall see, the
frontal lobe of the cerebrum is the "senior executive" in the
computation of individuality (ego) & volition, as well as (together with the
temporal lobe) abstract & creative thought. The cerebrum, especially the
frontal-temporal lobes, made
possible the explosion of meaning invoked by the Homo sapiens sapiens
(Cro-Magnon) and his highly developed neocortex.
the democratic neuron
Let us step back, and introduce the smallest operational unit of the nervous
system : the neuron. Only
members of the animal kingdom possess neurons. The neuronal kingdom appeared
around 700 million years ago, likely without axons and dendrites, secreting
electrical and chemical substances acting on other cells in a general,
non-specific way. Then a single, long transmission fiber emerged, the axon,
enabling the selective transmission of these messages to a second neuron.
Lastly, insulation was invented (myelin or fatty cells wrapping around the
axon, preventing leakage).
There are two broad types of cells in the nervous system :
neurons and glia ("glue"). There are about 130 billion neurons in the
human brain (of which ca. 20 billion constitute the cerebral neocortex) and
ca. 1.000
billion glia. The latter are thought to contribute to brain function mainly by
insulating, supporting and nourishing neighboring neurons.
Most neurons are in the range of 0.01 - 0.05 mm in diameter. The size of a
neuron is at or beyond the limit of what can be seen with the naked eye. The
central region containing the cell nucleus is the cell body (or soma),
whereas the tubes radiating away from the soma are called neurites, which are
either axons or dendrites. The soma usually gives rise to a single axon. Axons
act as "wires" carrying the output of the neurons (they travel
over great distances of a meter or more). Dendrites (extending rarely more than 2
mm in length) extend from the soma and taper to a fine point. They come in
contact with many axons and somata.
It was Cajal (1852 - 1934) who correctly argued the neurites are not continuous
from one cell to the next, communicating by contact. Earlier, Golgi (1843 -
1926),
the histologist who soaked brain tissue in a silver chromate solution, called
the Golgi stain, had maintained the opposite (both shared the Nobel Prize in
1906 but remained rivals). According to the latter, the neurites communicate by
way of fusion (a continuous network or reticulum similar to the arteries and
veins of the circulatory system). Cajal's neuron doctrine was finally proven
with the development of the electron microscope, with a resolution of about 0.1
nm, i.e. a million time better than the unaided eye, distinguishing two
points separated by more than one-tenth of a millimeter (0.1
mm).
"A handful of artificial neurons with a dozen-plus
connections can learn to tell the difference between apples and bananas. A dozen
with a few score connections can be taught to play Hexapawn or tic-tac-toe,
perfectly, in a few seconds. Fifty or so, with about 1200 connections, have in
reality done a better job of diagnosing heart attacks in an emergency room than
a team of expert cardiologists. A hundred neurons, with a few thousand
connections, can make money -a lot of money- in notoriously volatile markets. A
few hundred can mimic the development of human speech. Of what, then, is the 20
billion neuron, multiquadrillion connection human brain capable ?"
Satinover, J. : The Quantum Brain,
Wiley - New York, 2001, p.45.
The structure of the neuron is divided in soma & neurites (axons and
dendrites). The internal neuronal process gives rise to two fundamental states
of each living neuron : the neural membrane at rest or the action potential. The
external neuronal process is summarized by synaptic transmission.
A)
THE SOMA
The spherical cell body or soma of the typical neuron is 0.02 mm in diameter. A neuronal membrane separates the inside of the
neuron, composed of a salty, potassium-rich fluid or cytosol and a number of
enclosed structures or organelles, from the outside. The cell body of the neuron
contains the same organelles as those found in all animal cells, they are : the
nucleus and the cytoplasm (everything contained within the membrane excluding
the nucleus). The details of what happens in the cell falls outside our scope
(cf. gene expression, protein synthesis, Krebs-cycle in mitochondria, etc.).
The neuronal membrane (ca. 5 nm
thick and studded with proteins), enclosing cytoplasm & nucleus, excludes certain
substances floating in the fluid surrounding the neuron. It has been compared to a circus tent draped on an internal scaffolding or cytoskeleton. Its "bones"
contain elements dynamically regulated and in continuous motion.
B)
THE AXON
This is a structure unique to neurons. It is a highly specialized component for
the transfer of electrical current or nerve impulse over distances in the
nervous system. There is no protein synthesis in the axon, fed by the soma. It
is the different proteins in the axonal membrane enabling it to serve as
"wire". If an axon branches, it is generally at right angles (the
so-called axon
collaterals, which may be recurrent, i.e. return back to the original neuron).
The diameter of an axon ranges from less than 0.001 mm to about 0.0025 mm. The
speed of the nerve impulse is proportionate to the axonal diameter.
The beginning of an axon (the axon hillock) is differentiated from the axon
proper or middle part and from its axon terminal, terminal bouton at the end.
The terminal is the place where the axon comes in contact with other neurons or
cells, passing the nervous impulse on to them. This point of contact is called the
synapse ("to fasten together"). Axons may have many branches at their
ends (the terminal arbor) and each branch may form a synapse on dendrites or
cell bodies in the same region. Axons may form synapses at swollen regions or
"boutons en passant" along their middle part and terminate elsewhere.
When an axon makes synaptic contact with another cell it provides
innervation to it.
C)
THE DENDRITES
A dendrite ("tree") is a branch extending from the soma. The
dendrites of a single neuron or dendritic tree has a wide variety of shapes and
sizes used to classify neurons (in the cerebral neocortex there are two broad
classes : pyramidal neurons -with long axons- and stellate neurons -with short
axons-). The dendrites function as the antennae of the neuron and are covered
with thousands of synapses. The cytoplasm of dendrites resembles that of axons.
Some neurons have dendrites covered with dendritic spines regulating
synaptic input.
D)
THE SYNAPSE
At the synapse, the nerve impulse is transferred from one neuron to another. The
usual direction of this synaptic transmission, gives rise to presynaptic and
postsynaptic sides of the synaptic cleft. The presynapse is usually an axon
terminal and the postsynapse may be the dendrite or the soma of another neuron.
At most synapses, an electrical impulse traveling down the axon is converted in
its terminal into a chemical signal crossing the synaptic cleft. This signal is
converted on the postsynaptic membrane into an electrical impulse. This
chemical signal is a neurotransmitter stored and released from the synaptic
vesicles within the axon terminal. This typical
electrical-to-chemical-to-electrical transformation of information is one of
the great advantages of synaptic activity and makes possible many of the
brain's computations. Modifications of this process is involved in memory
and learning. Mental dysfunctions, nerve gas and psychoactive drugs are also
related to synaptic transmission, influencing the chemical balance.
In addition to anterograde transport (from soma to terminal), there is a
mechanism signalling the soma about changes in the metabolic needs of the axon
terminal. This is retrograde transport (from terminal to soma). The action
potential of a stimulated neurite does not jump on the membrane of the next
neuron, but only runs to the presynaptic side of the synaptic cleft. After a
short delay (cf. the transformation of the electrical impulse in a chemical
signal and back), this neuron reacts by lowering the potential of the neuronal
membrane (or axonal membrane). The level of depolarization increases with the
number of stimulating synapses (sum = a + b). Only if at the postsynaptic
membrane the depolarization-treshold is crossed will the next neuron transmit
the nerve impulse via its axon, etc.
E)
REST POTENTIAL
When a voltmeter measures the difference in electrical potential between the tip
of a microelectrode inside the neuron and a wire in the extracellular fluid, the
inside is ca. -65 Mv with respect to the outside. This is the restpotential or
equilibrium potential of the neuron (electrical potential, or voltage, is the
force exerted on a charged particle, reflecting the difference in charge between
anode and cathode). This electrical voltage is maintained by the neuron.
A sodium-potassium pump produces and maintains a large number of K+
ions across the neuronal membrane, leaving the inside of the neuron negatively
charged.
F)
ACTION POTENTIAL
When an electrical impulse hits a neuron by means of synapses (touching the soma
or the dendrites), the rest potential lowers a little (about +10 Mv) and when
the threshold of depolarization is reached, electrical couter-forces are
released. The neuron "fires" one action potential and this all-or-none
phenomenon moves with great speed from soma to axonal terminal (between 0.5 and
120 m/s, depending on the axonal diameter). When the potential enters the axon,
the permeability of the axonal membrane changes for a short period, while Na+
ions move to the inside of the axon. For a millisecond, the inside is positively
and the outside negatively charged. After this action potential or spike (+40
Mv), the rest potential becomes higher than before (ca. -80 Mv or
hyperpolarization), making the membrane refractive, i.e. uninfluenced by nerve
impulses. This allows the spike to move further forwards (direction axonal
terminal).
G) NEURONAL LOGIC
The neuron computes the logical operators "and", "or" and
"not" as follows :
Suppose neuron A, touched by two synapses B and C.
-
if B and C are both
stimulating A, A fires ;
-
if B or C (or both)
are stimulating A, A fires ;
-
C can only stimulate A if B
(inhibiting) is not stimulating A.
In reality, a neuron is fed by
hundreds of stimulating (yes) and inhibiting (no) nerve impulses. If the
stimulating impulses outweigh the inhibiting ones, the neuron "fires".
If otherwise, nothing happens and the neuron is "mute". This
impulse-frequency can modulate between 0 and about 250 action potentials per
second. The "democratic" principle is clear : majority
rules. Moreover, although individual neuronal operations have a relatively high
margin of error (compared to a computer), they are performed hundreds
of times simultaneously. This statistical method guarantees individual
errors weigh less on the final result. Another advantage is cortical plasticity
: groups of neurons adapting their activity after the destruction of another
group (cf. neuroplasticity).
the philogenesis of
the human brain
The embryo starts as a flat disks with three layers of cells : endoderm (the
internal organs), mesoderm (the skeleton and the muscles) and ectoderm (the
nervous system and the skin). Changes in the ectoderm give rise to the neural
plate, a flat sheet of cells.
In the neural plate a groove is formed, running from anterior (rostral) to
posterior (caudal). The walls of the groove or neural folds, move together and
fuse dorsally forming the neural tube. Out of the walls of this neural tube the
whole nervous system develops. When folding, some neural ectoderm is pinched off
lateral to the neural tube. All neurons with cell bodies in the PNS derive from
this so-called neural crest. The process from neural plate to neural tube or neurulation occurs about 22 days after conception.
The first step in the differentiation of the neural tube is the development at
its rostral end of three swellings called the primary vesicles, out of which the
whole brain emerges.
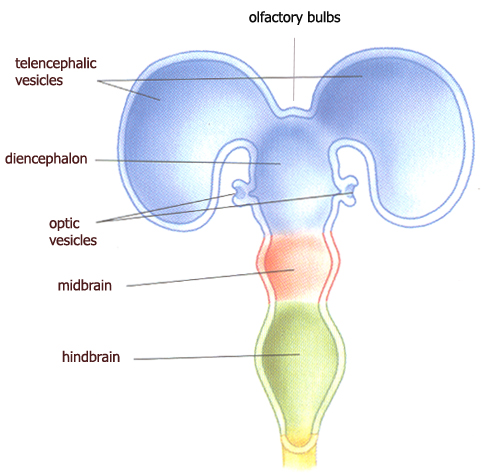
primary & secondary
vesicles of the neural tube
from
Bear, Connors &
Paradiso, 2001,
figure 7.10 p.182.
These swellings of the neural tube define the fundamental structure of the human
brain : forebrain, midbrain and hindbrain. The two cerebral hemispheres grow and
lie above and on either side of the diencephalon. The ventral-medial surfaces of
these hemispheres have fused with the lateral surfaces of the diencephalon.
Unlike the forebrain, the midbrain differentiates relatively little. From the
hindbrain, important structures emerged.
Out of the primary
vesicles, the following brainstructure emerges :
first primary vesicle : forebrain
* secondary olfactory vesicles : olfactory
bulbs (ventral surface of the hemispheres) - over the course of evolution, a group of primal
sensory neurons formed an internally located ganglion of like-minded cells
forming the olfactory bulbs, round projections part of the
forebrain - from the olfactory system, the
limbic system of the "old" mammalian brain evolved (ruled by the
amygdala) ;
* secondary optic vesicles : the two retina's & optic nerve are differentiated - the
major target of retinofugal projection is the superior colliculus
("little hill") of the tectum ("roof") of the midbrain ;
* secondary "telencephalic" vesicle : telencephalon
("endbrain") or cerebrum with its two cerebral
hemispheres, basal telencephalon (a pair of amygdala
and the hippocampus), the corpus callosum, cortical white
matter and the internal capsule ;
* secondary "diencephalic" vesicle : diencephalon ("between brain") : the thalamus and hypothalamus ; second primary vesicle : midbrain
: tectum and tegmentum ("floor"), with
the cerebral aqueduct in between ; third primary vesicle : hindbrain
: cerebellum, pons ("bridge") and medulla (oblongata).
4
Forebrain, midbrain &
hindbrain : structures & functions.
The brain dissected from the skull weighs
about 1.4 kg and resembles, in color and texture, a large piece of hard tofu.
Its common lateral view shows the "ram's horn" shape of the cerebrum
coming off the stalk of the brain stem. The different structures of the brain
emerge out of the fore-, mid- and hindbrain (the primary vesicles).
The
forebrain develops into the telencephalon (cerebrum, and hidden within it the
amygdala-hippocampal complex) and the diencephalon (thalamus & hypothalamus).
The midbrain into
tectum, cerebral aqueduct & tegmentum, and the hindbrain into cerebellum, pons
& medulla.
In doing so, this development calls into existence fluid-filled caverns and
canals inside the brain : the ventricular system. The organization of this
system is also used to understand how the mammalian brain is structured.
Although deviating from the general mammalian plan of the brain, in particular
due to the growth of the temporal lobes of the cerebrum, the embryonic
relationships between the ventricles and the surrounding brain still hold. One type of fluid runs in
the ventricular system (cerebrospinal fluid).
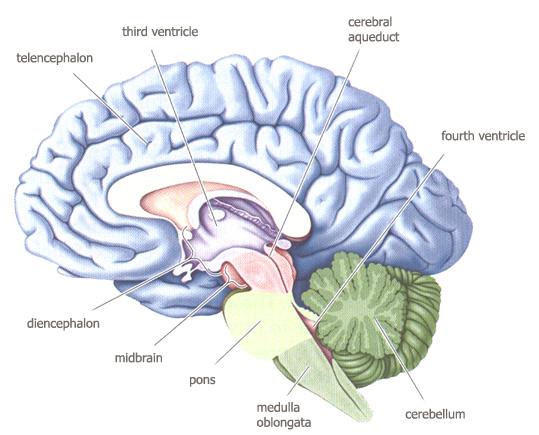
midsagittal view of the brain
from
Bear, Connors &
Paradiso, 2001,
figure 7.23 p.192.
According to the
ventricular system, the following
brainstructure emerges :
ventricles
(first & second) : cerebrum, 2 cerebral hemispheres ;
third ventricle
: in center of diencephalon ;
cerebral aqueduct
: in center of midbrain ;
fourth ventricle
: tube in hindbrain ;
spinal canal : in
spine.
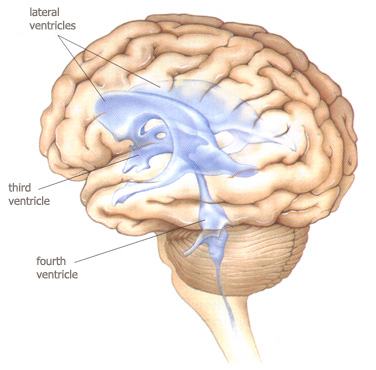
the ventricular system
from
Bear, Connors &
Paradiso, 2001,
figure 7.23 p.193.
FOREBRAIN
forebrain =
telencephalon + diencephalon
telencephalon = cerebrum
TELENCEPHALON
CEREBRUM
the lobes
The many convolutions on the surface of the human cerebrum (the sulci or
grooves, and the gyri or bumps) are its outstanding characteristic. They define the major difference between the
mammalian and the human brain. This increase in cortical surface is the
"distortion" of the human brain, and seat of the processing of
reasoning and cognition.
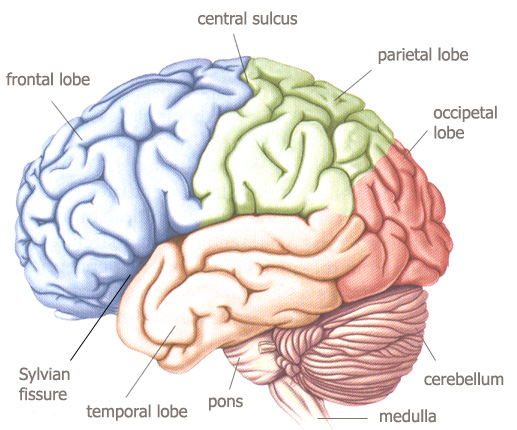
the cerebral lobes
from
Bear, Connors & Paradiso, 2001, p.207.
The cerebrum (measuring about 11 m²) is divided into
four lobes, situated underneath the corresponding bone of the skull :
-
the frontal bone of the
forehead covers the frontal lobe ;
-
the temporal bone (temple)
defines the temporal lobe ;
-
the parietal bone (caudal of
the central sulcus making the posterior border of the frontal lobe) covers
the parietal lobe ;
-
caudal to the parietal
lobe, under the occipital bone lies the occipital lobe.
basic structure
of the cerebrum
The cerebrum consists of gray and white cortical matter. Gray cortical matter is found in the cerebral neocortex (a thin sheet of
ca. 209 neurons
lying just underneath the surface of the cerebrum) as well as in the basal telencephalon ("basal" means buried deep
within).
The neocortex is that part of the cerebrum where sensations,
voluntary movement, learning, speach and cognition converge.
This neocortex (or set of neurons) shares several common features with all
vertebrate animals :
-
neurons are arranged in
layers or sheet, mostly parallel to the surface ;
-
the layer closest to the
surface is separated from the rest by a zone lacking neurons ;
-
at least one cell layer
contains pyramidal cells with large, apical dendrites extending upwards
& forming multiple branches ;
-
the cerebral neocortex has a
cytoarchitecture distinguishing it from the basal telencephalon.
At the beginning of the
twentieth century, the neuroanatomist Korbinian Brodmann (1868 - 1918) constructed a
cytoarchitectural map of the human cerebral neocortex. Each area having a common
cytoarchitecture is given a number. Later, it was put into evidence cortical
areas which look different perform different functions. Over the course of
evolution, the amount of cortex has changed, but not its basic structure.
Studying the differences between the neocortex and the primordial cortex of other
species (monkeys, cats, rabbits - cf.
Kaas, 2001),
reveals the human cortex consists of four types of "bark" :
-
primary sensory
areas : concerned with the reception of signals from the ascending
sensory pathways (all transiting via the thalamus) ;
-
secondary sensory areas :
assisting the primary sensory areas ;
-
motor
areas : concerned with the control of voluntary movement : these
receive inputs from thalamic nuclei relaying information from the basal
telencephalon and the cerebellum, and they send outputs to motor control
areas in the brain stem and spinal cord. This area is divided into primary
and secondary motor areas ;
association
areas : the remaining areas (particularly in the frontal and temporal
lobes) process the recent development of the primate cortex, namely the
ability to interprete behavior in terms of mental states.
In the association areas of the cortex
of Homo Sapiens sapiens,
sophisticated "human" processing occurs. These areas contain neurons
able to "associate" or "gather together" neural states from
various parts of the brain. Information from the sensory areas, memory systems
and the diencephalon (emotional states) is put together and integrated
in order to optimalize the possibilities of the nervous system and execute,
process, compute & enhance a conscious, aware observation of the world, as
well as each individual's efficient and good behavioral, emotional and cognitive
responses to it.
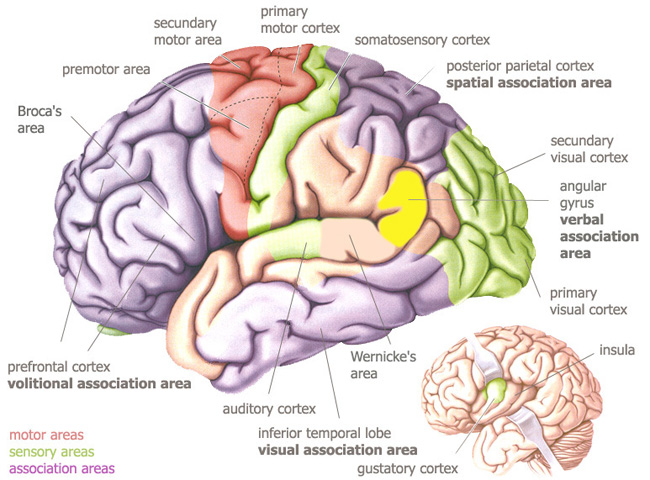 important areas of the
cerebrum
adapted from
Bear, Connors & Paradiso, 2001, p.208.
Four of these association areas have been discovered :
-
visual
association area : inferior temporal cortex : highest integration of
visual function & analysis - end station of a system of recognition of
specific and particular shapes and objects of interest, both cognitively as
well as emotionally - interconnected with the amygdala, hippocampus, limbic
system and the autonomous nervous system ; -
spatial
association area : posterior parietal cortex : highest integration of
analysis and integration of higher-order visual, auditory and somaesthetic
(touch & body position) information - three dimensional image of the
body in space - distinction between what is at arm's length (bodily sense)
and what is further away (the world) - some neurons motivate and guide hand
movements, including the grasping of objects within grasping distance ;
-
verbal
association area : angular gyrus, inferior parietal lobe : area of
the highest integration of sensory input, with rich interconnections with
all other association areas - processes abstract thought and their relation
to words (Wernicke & Broca in the left hemisphere) - conceptual
comparisons, ordering of opposites, naming of objects, higher logical
operations ;
-
volitional
association area (also : attention association area) : prefrontal cortex,
frontal lobes : receives fibers from all
sensory systems (vision, hearing, touch, taste, smell), but has few
connections with the primary sensory areas - very interconnected with the
limbic system (emotional responses), verbal and spatial association area
(conceptual thought and egocentic spatiality) - coordinates highly complex
movements and is the "seat of the will", for all goal-oriented
behaviors, actions and intentions - able to focus on important tasks through
redundancy (screening out superfluous input) - planning, imagining, deciding
and attention regulation throughout the cerebrum are computed here, but a
complete functional picture is far from clear.
the
"human" lobes
The frontal, temporal and
parietal lobes are essential in understanding how human consciousness is
processed (executed, computed) by the cerebrum of the Homo Sapiens sapiens.
A)
FRONTAL LOBE
The evolution of the frontal lobes made possible language (symbolization), tool
technology and artistic symbolism. They are the "senior executive" of
the brain (Passingham, 1993,
Fuster, 1989) and are primary in regard to all
aspects of imagination, creativity, speach, language (via Broca's area) and
symbolic thinking. In the frontal lobes, the coordination and regulation of
attention, individuality, memory and cortical activity is at hand. Intellectual, creative,
artistic, symbolic and cognitive processes get executed. They also subserve the
expression of melodic-emotional and vocabulary-rich grammatical speech.
B)
TEMPORAL LOBE
Temporal lobectomy on monkeys showed they suffered from "psychic
blindness", although able to see things, they did not understand with their
eyes what the objects were (cf. the visual attention area). The temporal lobes
contain the basal ganglia, discussed in the next paragraph.
C)
PARIETAL LOBE
Especially the inferior parietal lobe computes the most human of activities :
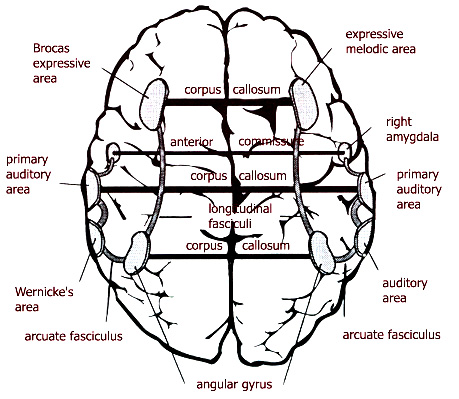
speech (cf. the verbal association area). Non-human mammals and hominoids lack angular gyri. These apes were limited to "hammering
with rocks, and throwing or manipulating leaves, sticks and twigs"
(Joseph, 2002, p.347). The control of temporal sequential hand movement,
manipulation of external objects & internal impressions are
particularly computed by the left angular gyrus of the inferior parietal lobe, unique to humans.
This gyrus is critically involved in "naming, word
finding, grammatical speech organization, and is in part an extention of and
links Wernicke's with Broca's area" (Joseph, 2002, p.357).
The
vocalization of meaningful sound (manipulating tools in a temporal, sequential
manner), multi-classification and multi-modality emerged with the Cro-Magnon.
Homo habilis, Homo erectus and Neanderthals "did not
posses the neurological sophistication for vocalizing complex human
language" (Joseph, 2002, p.358), i.e. angular gyri. The right
angular gyrus is crucial in performing artistic tasks. The Neanderthals had more
developed occipital and superior parietal lobes (larger in length and breadth), concerned with visual analysis and positioning the body in space
(cf. the importance of hunting for the archaic Homo sapiens).
special features of the
human cortical cerebrum
A)
CEREBRAL HEMISPHERES
The human cerebrum, unlike that of animals, is not symmetrical and each half
does not function in the same way. Duplication of effort is replaced by functional
specialization in need for cross-talk and collaboration, ensuring each
half can do "its thing" without interfering with what the other is
doing, while mutual assistance continues to move across the corpus callosum,
allowing for the computation of a unified consciousness and sense of "I" unique
to Homo
Sapiens sapiens.
The "human" reality expressed by the cerebrum is a dual-union, for
between both hemispheres information passes back and forth via the corpus
callosum, an axonal bridge between the two hemisphere, continuous with
cortical white matter (in humans, it is completed between 10 and 12 and
consists of a thick "cable" of 200 million nerve fibers). Cortical asymmetry is also suggested by the fact that 90%
of all Homo Sapiens sapiens have a preference for the right side,
indicative of a dominant left hemisphere.
Following functional differences
prevail :
after
Joseph, 1993, p.44
|
Left
Hemisphere |
Right
Hemisphere |
| linguistic |
kinesthetic |
| propositional |
visual |
| discrete |
diffuse |
| analytical |
synthetical |
| verbal |
visuospatial |
discursive
(logical) |
presentational
(melodic) |
| digital |
analogical |
| specific
features |
broad features |
| deliberate |
analogical |
This functional asymmetry does
not impair the cerebrum to function as a whole. Thanks to the corpus callosum,
left and right cerebral approaches may be balanced. Thus, cerebral activity can
be summarized under the heading of "the union of opposites", just as
in the image of the two scales of the balance. Synchronization of both
hemispheres, which are in a constant state of flux, is one of the keys to a balanced
approach of life and an healthy response to it.
B)
LATERALIZATION
Is the "dominance" of the left hemisphere over the right, i.e. left
lateralization, a normal phenomenon associated with the processing of human
consciousness by the cerebrum, or rather a sign of a disrupted capacity to
process a unified consciousness ? Biofeedback confirms the importance of a
balanced (symmetrical, synchronized) cerebral activity. The notion the
right hemisphere is somehow inferior, minor or animalistic has been
relinquished. Split-brain patients demonstrate how the "silent" hemisphere has a
mind of its own ! So, if one hemisphere dominates the other, cerebral activity
will be less interesting than in a balanced, two-way cerebral momentum. Each
hemisphere performs different parts of an integrated computation. The plasticity
of this performance is as extraordinary, as the role of the corpus callosum is
essential. The category major/minor is thus interchangable.
"It has now been clearly demonstrated that the right
hemisphere is dominant over the left in the analysis of geometric and
visual-space, the perception of depth, distance, direction, shape, orientation,
position, perspective and figure-ground, the detection of complex and hidden
figures, the performance of visual closure, gestalt formation, and the ability
to infer the total stimulus configuration from incomplete information, route
finding and maze learning, localizing targets in space, drawing and copying
complex geometric-like figures and performing constructional tasks, block
designs and puzzles."
Joseph, 2002, p.354.
C)
INSULA
The insula ("island of Reil") is a piece of cerebral cortex (the
gustatory) bordering and separating the temporal and frontal lobes.
D)
COGNITIVE OPERATORS
Cognitive operators are the mind's most general analytical functions or primary
functional components (d'Aquili & Newberg, 1999,
p.50). They differ from cognitive modules (Pinker, 1999), which represent more
specific localized functions. Operators are analogous to those used in
mathematics, relating two mathematical elements (addition, substraction,
multiplication and division). The seven cognitive operators proposed by d'Aquili
& Newberg (holistic, reductionist, causal, abstractive, binary,
quantitative, emotional value) have been criticized by
Atran
(2002, pp.183-184). The reader is referred to the
epilogue.
FOREBRAIN DEEP
forebrain =
telencephalon + diencephalon
cerebrum = gray matter + white matter
gray matter = neocortex + basal telencephalon
BASAL TELENCEPHALON
The neuronal structures underneath the cerebral neocortex, or
subcortical structures of the "deep" or "basal" telencephalon,
interconnect the
neocortex with the diencephalon.
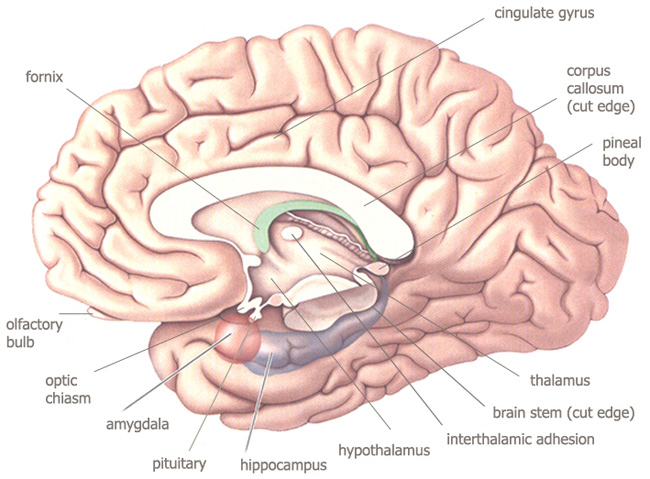
fore- & midbrain : brain
stem and cerebellum removed
adapted from
Bear, Connors & Paradiso, 2001, p.211.
Deep within the telencephalon, two important structures emerged : the
hippocampus ("seahorse") and the amygdala ("almond").
Together, they constitute the amygdala-hippocampal complex.
HIPPOCAMPUS, the archive-keeper and emotional equilibrator
The hippocampus is a piece of cortex situated in the temporal lobe with only a
single cell layer connected with the olfactory cortex (for continuous with the
olfactory bulb). It is influenced by the activity of the amygdala and both often
act together in a complementary way (emotional interesting sensoric input linked
with images, memory & learning). The hippocampus has regulatory effect on
the thalamus. It may block (with or without the thalamus) sensory input to the
neocortex and regulate the autonomous nervous system by maintaining emotional
equilibrium. As such, the hippocampus does not process the generation of
emotional states, but memorizes them. Although the amygdala plays as large a
part in memory as the hippocampus, the latter serves to both consolidate and
construct memory.
AMYGDALA, security agent and emotional regulator
The amygdala are a complex of nuclei situated in the pole of the temporal lobe,
just below the neocortex on the medial side, on both sides of the cerebrum. Afferents to the amygdala come from
all lobes of the brain. Information from all sensory systems feeds into them,
each having a different projection pattern to these nuclei. The amygdala are also
connected with the hypothalamus and may activate it, although they modulate &
control its drives. They may act at the behest of the hypothalamus and mediate all
higher-order emotional functions (spiritual included), have a surveillance
function and assign emotional value to stimuli.
The amygdala may
overwhelm the cortex and, in the mammalian brain, they wrested the master control of emotion from the
hypothalamus (the "master controller" of the reptilian brain - cf.
infra). The amygdala are the master controller of the emotional brain.
Both the hippocampus and the amygdala are part of the temporal lobe, and play an
important role in connecting the two parts of the forebrain : the telencephalon
(the cerebrum, especially the neocortex) and the diencephalon (thalamus and hypothalamus).
DIENCEPHALON
The diencephalon differentiates into thalamus
and hypothalamus. These structures are part of a larger whole, called "the
limbic system".
LIMBIC SYSTEM
In 1878, the French neurologist Broca (1824 - 1880) discovered mammals possess a
group of
distinctly different
cortical areas.
This collection of neurons formed a ring or border ("limbus") around
the brain stem. In his definition, the "limbic cortex" consists of the cortical
cortex
around the corpus callosum (especially the cingulate gyrus) and on the medial
surface of the temporal lobe (including the hippocampus). By the 1930s, it
became clear a number of these "limbic" structures were involved in
emotion. James
Papez proposed an "emotion system" linking the
neocortex with the hypothalamus (forming the Papez circuit), each element
being connected to another by a major fiber tract (like the fornix,
"arch", a major
bundle of axons leaving the hippocampus).
To physically experience emotion, the neocortex is critical, but to express
it, the hypothalamus is imperative. The cingulate cortex projects down to the
hippocampus and the latter down to the hypothalamus by way of the fornix.
Upward hypothalamic effects reach the neocortex via a relay in the nuclei of
the anterior thalamus.
The Papez circuit is bi-directional :
Because of the similarity
between the elements composing both the Papez circuit and Broca's limbic lobe,
this group of structures is referred to as the lymbic system (although Broca's
notion did not focus on emotion). The limbic system is part of the triune brain
(cf. infra).
THALAMUS, the gateway
The thalamus ("inner chamber") is nestled deep inside the
forebrain (dorsal part of the diencephalon) and highly interconnected with the
neocortex, i.e. the upper walls of the third ventricle. The spinothalamic pathway is the major
route by which afferents (registering for example pain or temperature) ascent to
the cerebral cortex.
The thalamus is the gate through which information
carried by sensoric axons enters the CNS. Here, these
afferents are pre-processed to branch out to the relevant cortical areas and the
limbic system. It can inhibit the flow to and fro the neocortex, integrates a
variety of inputs and translates them into a format the neocortex can read. So
the thalamus projects into various cortical areas.
HYPOTHALAMUS, the master controller of the ANS
Philogenetically, the hypothalamus is the most ancient of diencephalic structures. It is situated near the upper end of the brain stem. The inner
part of this structure (not larger than a pea) is an extension of the
quiescent system, involved with the baseline function of the organism (the
parasympathical branch of the ANS) and a subjective sense of peace and
calmness ("rest & digest" response). In contrast, the outer edge is the extension of the arousal system,
the fight-or-flight response (terror, rage) as well as the experience of
extreme positive emotions (from pleasure to bliss). These responses are
stimulus bound and die off when the stimulus is removed. Hence, this nucleus
is the master control for the ANS (integrates its actions), able to affect every organ or part of the
body. It links its operations (arousal or relaxation) to the higher cortical
functions. Through it, the latter are able to co-regulate body functions. The
hypothalamus also controls aggression, sex and survival-behaviors. It
regulates many hormonal systems, including reproductive hormones, the thyroid,
growth hormone and moderates hunger, thirst and body temperature.
MIDBRAIN
forebrain =
telencephalon + diencephalon
midbrain = tectum + cerebral aqueduct + tegmentum
The midbrain is only a narrow
CSF-filled space, the cerebral aqueduct, with a tectum ("roof") and a
tegmentum ("floor"). Although simple, the functions of the midbrain
are diverse : a conduit for information passing from the spinal cord to the
forebrain and back, neurons contributing to sensory systems & the control
of moment, axons descending from the cortex to the brain stem and spinal cord
(corticospinal tract), related to the voluntary control of movement.
The superior
tectum receives direct input from the eyes. Axons supplying the eye muscles
start there (cf. eye-movements). The inferior tectum receives sensory
information from the ears and relays auditory data to the thalamus. Of the
motoric afferents passing through the midbrain, 90% (or 20 million axons in the
human brain) synapse on neurons in the pons. These pontine cells relay this to
the cerebellum on the opposite side. The pons is a massive switchboard
connecting the cerebral cortex to the cerebellum.
HINDBRAIN
forebrain =
telencephalon + diencephalon
midbrain = tectum + cerebral aqueduct + tegmentum
brainstem = midbrain + pons + medulla
hindbrain = cerebellum + pons + medulla
Like the midbrain, the hindbrain is also a conduit of information passing from the
forebrain to the spinal cord and back. Its neurons contribute to the processing
of sensory information, the control of voluntary movement and the regulation of
the ANS.
To regulate the overall activity-control of the brain, neurons with a
particularly widespread pattern of axons are called in. They use particular
neurotransmitters and have widely dispersed, diffuse connections, modulating vast
assemblies of postsynaptic neurons, such as the cortex, the thalamus etc.,
causing them to be more or less synchronously active (more or less excitable).
These very diffuse modulatory systems share common features :
-
they consist of a small set
of neurons (several thousand) ;
-
they arise from the core of
the brain, most of them from the brainstem ;
-
each neuron has an axon contacting more than 100.000 postsynaptic neurons across the brain ;
-
their axons have a transmitter
which is not confined to the vincinity of the cleft, but diffuse to many
neurons.
THE RETICULAR FORMATION, the
gate keeper, the filter, the master modulator
The ca.12.000 neurons of the tiny locus coeruleus ("blue spot" because
of the pigment of its cells), of which we have two, one on each side, have axons fanning out to innervate about every part of the brain. One neuron can
make more than 250.000 synapses and can have one axon branch in the cerebral
cortex and another in the cerebellar cortex. On either midline of the brain stem
lie the nine raphne ("seam", "ridge") nuclei. Each nucleus
projects to different regions in the brain. Together with the locus coeruleus,
the ascending reticular activating system (ARAS) is constituted. This is
the "core" of the brain stem, involved in the regulation of attention,
arousal, sleep-wake cycles and general metabolism of the brain. The ARAS system
arouses and awakes the prefrontal cortex. Raphne neurons control the sleep-wake
cycles and well as the different stages of sleep.
a diffuse modulatory system :
the locus coeruleus
adapted from
Bear, Connors & Paradiso, 2001, p.515-518.
In this region of their brain, humans assemble a "set of
instructions" for purposes of long-term storage (MacLean, 1976),
with dreams playing an important part in this process & integration in general. The general immediate assessment of place, time & person
followed by ongoing evaluation is the automatic, "instinctual" orienting response (also acting as a
filter to block certain inputs) processed here.
|
REGION
OF CNS |
EXECUTIVE
FUNCTION |
| neocortex |
higher order
cognitive, affective and motoric processes |
| cerebral
hemispheres |
concert of
analytical and synthetical higher-order approaches |
| prefrontal
cortex |
senior
executive of all
higher order functions |
| cingulate
gyrus |
willful acts,
intent |
| angular gyrus |
senior
language controls |
|
threshold
between cortical & subcortical structures |
| hippocampus |
memory
consolidation
and emotional equilibration |
| amygdala |
security and
emotional regulation |
| thalamus |
universal
gateway
integration & projection |
| hypothalamus |
master
controller
of the ANS |
| limbic system |
overall
emotional circuit |
| ARAS |
filter and
master modulator |
| ANS |
basal
neurophysical activity |
cerebellum
The cerebellar cortex is an important movement control center, receiving massive
axonal inputs from the spinal cord and the pons (position of body in space).
Goals of intented movements originating in the prefrontal cortex are relayed via
the pons. The cerebellum compares and calculates the sequences of muscle
contractions required.
5
The triune brain : reptilian, mammalian and human.
Neuroscientist
MacLean (1970,
1 978, 1990) advanced the concept of the triune
brain. In the brain as a whole, he identified a three-tiered structure, called
"reptilian", "mammalian" and "neocortical". This
division has been used by neurophilosophy and contemplative science to explore the meeting between
religion and neuroscience (Albright & Ashbrook, 2001). MacLean and other
researchers have observed animals whose activities depend on each of these
"brains". In this way, various functions have been attributed to each
of these three parts of the brain, characterized by a different structure and
chemistry, yet extensively interconnected.
-
the
reptilian brain : brain stem (midbrain, pons, medulla), midbrain,
hypothalamus ;
-
the
mammalian brain : thalamus, hippocampus, amygdala ;
-
the
human brain : neocortex of cerebral hemispheres of cerebrum, angular
gyrus.
Compare this with the anatomical
division of the brain in fore-, mid-, and hindbrain :
Reptilian brain =
hindbrain
+ midbrain + hypothalamus (the "oldest" master controller)
;
Mammalian brain
=
higher
thalamic diencephalon + basal telencephalon (hippocampus and amygdala, the "old" senior
executive) + cingulate
cortex (relay to cortex)
;
Human brain = forebrain,
neocortex (the "new" senior executive, in particular the prefrontal
cortex).
In this context, stress the difference between mental states of which the
subject of experience is conscious (processed by the neocortex, in particular
the prefrontal lobes) and those remaining unconscious (processed by the
dominated hemisphere, the limbic system and the brainstem). The former can be
studied directly (for direct conscious feedback is possible), whereas the latter
is made evident in the emotional coloration of information and the regulation of
the overall wakefullness of the brain. The present neurophilosophical
investigation aims at a partial, incomplete & provisional understanding of the
nervous system as a whole, i.e. peripheral nervous system included. The
studies of Maclean accommodate such an approach.
A) THE REPTILIAN BRAIN
The behaviors of reptiles are ritualistic, conservative and
"programmed", with no playfulness, joy or sadness. There are 4 main
types of reptile display or rudimentary communication :
ying members of
the same species ;
courtship
: signals to attract the attention of the opposite sex ;
territorial
challenge : instinctual attempt at deception to fend off invaders or
attackers ;
submissive/dominance
: strict hierarchy and ritual submission allows the group to survive.
In the reptilian brain of
non-reptiles, the
territorial imperative (cause of war) is still at work, in life support and self-protection. In reptiles, the latter is often
realized through deception, imitation & secrecy (cf. the treacherous
serpent, the violent crocodile, the cowardly cameleon, the slow tortoise). The
responsive (not reactive) nature of this brain, makes it suitable for guarding,
patrolling and vigilance, which are bound to routine, precedent and ritual. The
conservatism of this brain is stabilizing, and routinizing saves energy.
However, a failure to adapt, is characteristic of it. In the brain stem,
addiction & deep memory storage through dreams are processed. The reptilian
brain houses the more violent, aggressive -even murderous- expressions of
impersonal sexuality, expressing a vertical, hierarchical, dominance/submission
signal communication.
the reptilian brain has two
major neurological structures :
the ARAS and the hypothalamus
This earliest brain and its "automatic pilot" are anchored in the
embodied, the concrete. It has no emotions and no language, except signals.
Traditional religious, philosophical and cultural systems (computed in the
prefrontal cortex) devise rituals to exorcize (banish, cast out or at best tame)
the troublesome and unwholesome urges & drives of the reptilian brain and
keep one "pure". The whole strategy is dualistic. By
dividing and ruling, theologies do not assist in the integration of the
triune brain, which, on the lowest level, implies a rhythmical ride "on the
back of the crocodile", knowing the precise momentum to co-opt "the
Monster" and to create a win-win situation instead of making us captive of
the division between the "law of God" and "the law of sin that
dwells in my members" (cf. Paul, Letter to the Romans, 7:21-24).
B)
THE MAMMALIAN BRAIN
Reptiles fight without
excitement, show no panic at being prey and display no hot-blooded couplings,
just ritualized (at time violent) behavior. Mammals, to a greater or lesser
degree, exibit anger in competition, cravings for food and lustful drives to
mates.
Emotion is the motivating & mobilizing source of empathy and memory and
unites relatedness and nurturance. In reptiles, the limbic structures around the
brain stem are present in abbreviated form only. They are found in all mammals and specialize in attaching emotional labels to the incoming and outgoing
streams of information. This brain is able to trigger emotions related to
eating, mating, fighting (food, mates), the care of the young, play &
community. Relating and nurturing are the essential feature of this brain (Albright &
Ashbrook,
2001, pp.85-86).
the mammalian brain has
three major neurological structures :
the thalamus, the hippocampus and the amygdala
Emotion labels this-or-that as something which matters to us.
Emotion adds colour & affect to mental states. Emotion
enables permanent storage (in the brain stem), suggestive of the ongoing
activities between the three levels of the living brain, computing mind. While emotional memory is formed
and its individual, emotional aspects stored in the amygdala, cognitive, visual
and contextual variables are stored (recalled) by the hippocampus (Gloor,
1997).
C)
THE HUMAN BRAIN
To be able to compute all higher-order operations is the "nominal"
mode of working of the cerebrum and its specific, bi-modal approach : two
hemispheres processing one integrated cerebral activity from two different
angles. Abstract thoughts can be thoroughly computed after the axonal bridge between
both, the corpus callosum is completed (cf. Piaget's
"formal-operatoric phase" after the age of 10).
Contrary to the reptilian, mammalian and all other cortical brains on Earth, the neocortex of
Homo Sapiens sapiens is exceptional in size, wiring & function. Of all mammals,
humans have the most "uncomitted cortex" at birth (Penfield, 1975),
i.e. fewer neurons with, in their hardware, instinctual patterns
built-in. This implies
the human brain is made for organic neuroplasticity (the more difficult a task, the
more cells process it) and also has great ability to learn and
individualize.
the human brain has one major neurological
structure :
the neocortex
The bi-modality of the human
brain is horizontal & vertical.
On the horizontal horizon, there is the joint project of the two cerebral
hemispheres : cerebral activity is called to be an integration of a duality.
This is accepting the difference while opening up as many neuronal alleys
between the hemispheres (cf. the "concordia discors" of thought in
Criticosynthesis, 2008).
Vertically, the neocortex (or upper telencephalon) and the basal
telencephalon perform different tasks Although 95% of right-handed people have
left-hemisphere dominance for language, only 18.8% of left-handed people have
right-hemisphere dominance for language function.
The basal telencephalon is part of the limbic
system. It is essential in the relay of information down from and up to the
neocortex and adds "emotional color" to what comes in and goes out.
Especially the amygdala play a crucial role in this, while the
association of memory & emotion is noteworthy.
-
left
hemisphere/neocortex : higher order
verbal operations ;
-
left
hemisphere/basal telencephalon : emotion/word associations, digital
memory ;
-
right
hemisphere/neocortex : higher order visuospatial operations ;
-
right
hemisphere/basal telencephalon : emotion/imaginal sensations,
visual memory.
|

